4.1 人亚科原人的进化:从 800 万年前到 20 万年前
04-12Ctrl+D 收藏本站
上一章我们提到了赤道非洲地区栖居树上的类人猿(great apes)。到了大约 800 万年前,除了亚洲猩猩(orangutan)之外,早期类人猿在非洲之外的世界其他地区都灭绝了。
人类是从类人猿的一支进化而来的。长期以来,我们都不清楚这支类人猿是大猩猩(gorilla)还是黑猩猩(chimpanzee),直到遗传研究表明,人类更接近黑猩猩,人类与黑猩猩的基因相似度达 98.5%,从而确定了它们是人类最近的亲属。现代黑猩猩和现代人类是从一个共同祖先进化而来,大约 800 万年前到 500 万年前,我们人类才分离出来。通过估算两个物种基因出现差异所需时间(基于遗传变化的大致速度),就可以得出这个结论。
与黑猩猩从共同祖先分离出来的那支猿人的各个种(其中也包括人类),我们称之为人亚科原人(hominine)。人亚科原人这个词可能有点让人困惑,因为直到最近被使用的词汇是原人(hominid)。不过,与以前所想的一样,遗传研究已经表明,黑猩猩和倭黑猩猩(bonobo,黑猩猩的另一个种)更接近人类,而不是更接近大猩猩。因此,分类体系需要做出调整,不过,科学家还没有在如何调整上取得一致。这样,原人的含义就取决于所使用的是哪一种分类体系。所有生物分类学都使用了相同的总体层次,比如界、纲、科(参见词汇表),然而,在亚群的内容上,生物学家出现了分歧。我们使用的体系如表 4.1,其中所有类人猿都被纳入一个总科;同属一科的类人动物(hominoids)——包括大猩猩、黑猩猩、倭黑猩猩以及人类——称为人科(hominids);亚科中人类的所有后代被称为人亚科(homininae)或人亚科原人。人们就这些分类应当如何排列争论不休。
以下关于我们物种的分类,展现了我们人类慢慢出现的各种特征。
人类属于
·真核生物总界(由真核细胞构成);
·动物界(非植物或真菌)
·脊索动物门(具有脊骨的动物)
·哺乳纲(哺乳幼仔的脊索类动物)
·灵长目(大型的、栖息树上的哺乳动物)
·人猿总科(人类和所有类人猿,包括黑猩猩、倭黑猩猩、大猩猩、长臂猿和猩猩)
·人科(包括大猩猩、黑猩猩、倭黑猩猩和人类)
·人亚科(两足猿〔双脚直立姿态〕,与黑猩猩分离以后人类的所有种)
·人属(双足猿,大脑大于 800 立方厘米)
·智人种(解剖学上的现代人,人亚科唯一幸存的种群)
人类进化的证据类型
最近 60 年,我们在理解人类进化方面取得了巨大进展,因为测年技术得到极大改进,同时也存在其他类型的证据。我们简要描述来自以下四个领域的证据:史前考古学(研究骨骼化石和石器工具)、灵长类学(研究现代灵长类动物)、遗传学(研究基因)以及气候学(研究气候变化)。
化石骨骼与石器工具
19 世纪中期,欧洲科学家开始研究人亚科原人的化石骨骼和石器工具。1850 年,西西里和法国北部发现的石器工具的绘图得以公布。1857 年,尼安德特人(Neandertals)的最早化石在德国尼安德山谷被发现,不过,第一批古代人类骨骼(从解剖学而言,也是现代人类骨骼)是在 1868 年发现于法国莱塞济附近克罗马农山洞,因此,它们被称为克罗马农人(CroMagnon)。
20 世纪早期,绝大多数欧洲科学家认为,直到大约 6 万年前,智人才在欧洲进化出来。二战之后,史前考古学家开始在非洲寻找骨骼;其中著名的有路易·利基(Louis Leakey,1903-1972)与他的妻子玛丽·利基(Mary Leakey,1913-1996)以及儿子理查德·利基(Richard Leakey,1944-),他们在东非大裂谷地区坦桑尼亚奥杜韦峡谷(the Olduvai Gorge)从事考察工作(参见地图 4.1 和第 2 章)。理查德·利基扩大了他的考察活动,1967 年,身在埃塞俄比亚的他发现,路易·利基所说的骨骼,正是非洲最早的智人骨骼。到 20 世纪 90 年代早期,许多考古学家承认,智人肯定是在非洲进化出来的。达尔文在他的著作《人类的由来》(1863)中就做出了类似推论,他的依据在于,非洲存在我们的近亲,即大猩猩和黑猩猩。
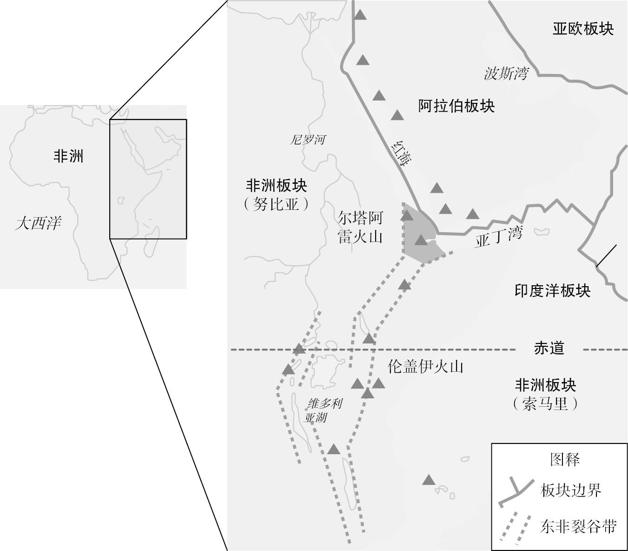
地图 4.1 东非大裂谷的研究地点。
东非大裂谷是由非洲大陆板块的一条裂缝形成;裂谷东部正在分离,有朝一日会漂浮在印度洋上。阿法尔三角带处于三个相互分离的板块——阿拉伯板块和非洲板块的两块碎片(努比亚和索马里)——的结合部。大裂谷导致了历史上很活跃的火山(三角地区),它们洒落的火山灰掩盖了尸体,也保存了骨骼
非洲还出现了比智人更早的人亚科证据。1960-1963 年间,利基一家找到了一些遗迹,他们称之为能人(homo habilis, 意指“巧手人”,因为它们似乎制造了石器工具)。能人生活在大约 250 万年前到 175 万年前。1974 年,唐纳德·约翰松(Donald Johanson,1943-)在埃塞俄比亚的阿法尔三角地带(the Afar Triangle)找到了更古老的骨骼,他把其中一幅不太完整的著名骨架称为露西(Lucy),露西大约生活在 320 万年前。两年之后,玛丽·利基在坦桑尼亚利特里(Laetoli)的火山灰中发现了人亚科原人的足印,年代大约为 350 万年前。1994-2004 年间,8 种新的早期人亚科原人种被发现,这 10 年也成为人类古生物学成果丰硕的时期之一。
化石遗迹能够告诉我们大量关于我们祖先的信息。这类化石不但可以告诉我们它们的形体以及它们如何行走和移动,也可以告诉我们它们大脑的容量、居住的环境,甚至它们是素食者还是肉食者,如果我们仔细研究它们的牙齿的话。
现代灵长类动物
对现代灵长类动物的研究,能够为我们提供间接证据来了解我们祖先可能是如何生活的,尽管在这类证据能够告诉我们多少关于祖先的信息方面,人们还存在争议。直到 20 世纪 60 年代之前,我们对野生灵长类动物知之甚少;大多数专家只是在动物园进行研究活动。此后 20 年,灵长类动物学兴盛起来,尽管受到重重阻碍。热带雨林正在消失,随之消亡的,还有雨林中大量的猴子和类人猿。研究者常常发现,他们不但很难研究自己的研究对象,同样也很难保护它们。
简·古多尔(Jane Goodall,1934-)最初与利基一起工作,1960 年,她前往坦桑尼亚坦噶尼喀湖(Lake Tanganyika)地区的贡贝河野生动物保护区(Gombe Stream Game Reserve,目前已经成为国家公园),并且与黑猩猩住在一起,由此开始了对野生黑猩猩的开拓性研究。她的报告震惊了世人,因为她发现黑猩猩使用工具:用棍子捕集白蚁窝中的白蚁、用石头砸开果实、用树叶擦掉粪便。让古多尔感到惊讶的是,医学研究者早就认识到了黑猩猩和人类在生理学上的相似性——比如,我们能够相互输血——但是,他们却忽视了两者之间在情感和社会方面的密切相似。通过揭示人类行为多么符合动物世界的行为,古多尔引起人们态度的变化。1965 年,西田利贞(Toshisada Nishida)开始研究坦桑尼亚马哈尔山的黑猩猩,他也做出了重要贡献。
基因比较
20 世纪 60 年代以来,一种新证据出现,即科学家学会比较不同物种的基因。我们由第 3 章得知,当两个物种彼此分离时,中性突变(neutral mutations)就会在每一支中累积起来。中性突变是指基因中非编码 DNA 的改变,这部分 DNA 不会提供制造蛋白质的指令,也不会被自然选择清除。目前为止,我们还不能理解基因组中非编码的、中性的或沉默的基因的功能。在每一个物种中,中性突变的积累速度非常稳定,因此,中性突变的数量可以揭示物种有多古老。研究者通过计算两个现代物种 DNA 序列之间的差异,就可以估算出它们何时从共同祖先那里发生分离,或者他们通过计算一个基因中中性突变的数量,就可以估计出那个基因盛行于何时。
1967 年,基因测年首次被用来估算人类与非洲类人猿的共同祖先,得出的结论是,这一祖先生活在大约 700 万年前,大多数古人类学家表示强烈反对,他们认为,更有可能的年代是 2000 万年前到 1500 万年前。然而,进一步的研究不断表明,人类与类人猿的分离时间发生在 800 万年前到 500 万年前,大多数古人类学家现在已经接受了这些年代。
气候变化
20 世纪早期,科学家们认为,全球气候在几千年时间里只会发生缓慢变化。20 世纪 50 年代,少数科学家利用碳-14 测年获得的证据表明,过去一些气候变化只发生在几千年时段。20 世纪 80 年代和 90 年代,有证据显示,全球气候可能在一个世纪甚至短短十年之内就发生改变。这种证据主要来自对大块冰原的冰核钻探研究——深入 3.2 千米。钻探取样的一块典型冰柱,直径大约 10 厘米,长度为 30.5 厘米,这样,用强大的显微镜就能够分析冰块的年层。每一层都含有形成时期的微型空气泡,通过分析空气泡,就能够确定当时大气的构成以及地球表面的平均温度。
气候变化的证据不但来自冰核样品,也来自海底沉积物的样品。海底样品讲述了一个更久远的故事,远至 100 万年前,而冰核样品则提供了一幅更详细的、每一年的图景。通过使用来自格陵兰岛冰核的高分辨率证据,气候学家差不多能够以十年为单位标绘出过去 80 万年的气候变化,能够以月为单位标绘出过去 3000 年的气候变化。对大量存在的耐寒花粉进行研究,也有助于我们理解气候变化。因为不同植物物种的花粉不同,由此可以重建整个植物生态系统。
在过去 70 万年,地球的气候随着每 10 万年一次的冰期和其间夹杂的长约 1 万年的短暂的温暖期(或“间冰期”)而波动。冰核证据表明,在最后一次冰期的最后 1000 年,暖冷频繁更替,剧烈的变化会在数年或数十年发生(参见图 4.1)。如果这个模式适用于今天的话,那么,最近 1 万年的温暖期正在结束,我们会再次进入一个冰期。这些模式表明,人类是在气候很不稳定的一段时期进化的,这种不稳定性或许有利于适应能力很强的物种的进化。
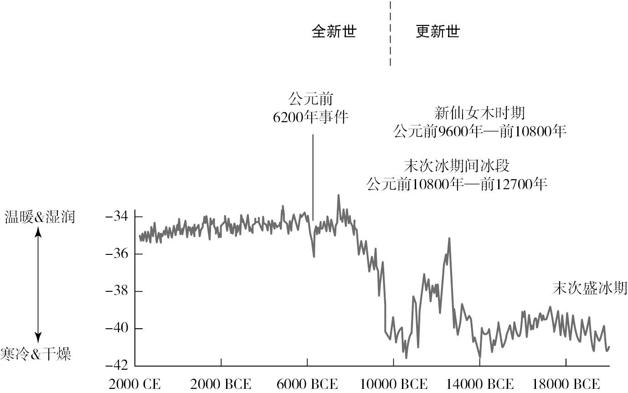
图 4.1 公元前 2 万年至今的全球气温。
这幅图表的依据,基于取自格陵兰岛的一块冰核之化学成分的变化。被检测的是氧的两个同位素氧 16 和氧 18 的相对存量。请留意,最近 1 万年与此前 1 万年相比,气候更暖和也更稳定
在过去一代人的时间里,科学家已经学会了把气候的改变与地球公转轨道的变化联系在一起。轨道的变化影响了每年不同时刻太阳辐射抵达地球的总量。这种变化有周期性,被称为米兰科维奇循环(Milankovitch cycles),因为正是塞尔维亚天文学家米卢廷·米兰科维奇(Milutin Milankovitch,1879-1958)发现了这个循环。地球轨道有三种不同变化。第一种是地轴所指方向的改变,即地轴的摆动,变化周期大约为 21000 年。第二种是地轴倾斜度的变化,倾斜角度介于 22.1 度到 24.5 度之间,变化周期为 41000 年。第三种是地球公转轨道不是一个完美的圆,这是地球附近其他行星引力作用的结果,变化周期为 10 万年和 40 万年。自大约 3500 万年前以来,这些米兰科维奇循环就构成了我们的行星时钟;它们的影响现在还可以在岩层中发现。
人亚科原人发展的不同时期
一想到人类与黑猩猩有共同祖先并由此进化而来,是不是觉得很别扭呢?许多人确实如此感觉,可能他们没有把推算出来的进化所需的 700 万年,即大致 28 万代人的时间考虑在内。
表 4.2 概括了现代黑猩猩与现代人类之间的主要差异。我们后面会探讨几百万年时间内发生的这些变化,我们分三个时期——雨林、稀树热带草原、灌丛热带草原进行讨论,每个时期都出现了标志性的环境变化。
黑猩猩
人类
指关节拄地行走
两足动物
脑容量小(大约 1/3)
脑容量大
牙齿、颚、嘴较大
牙齿、颚、嘴较小
毛发黑,肤色浅
毛发少,肤色深(现在多变)
合群
更合群
喉位高
喉位低
无须协助的独自生育
协助下的社会分娩
男性体型比女性大 25%~30%
男性体型比女性大 15%~20%
男女等级制
配对结合
独自进食
群体进食
不使用火(生吃)
用火(熟食)
简单工具
复杂工具
发声与姿势
完全的句法表达
24 对染色体
23 对染色体
雨林
现代黑猩猩生活在赤道热带雨林,即此前所说的丛林(jungle,印地语森林的意思)。雨林炎热、潮湿,布满了快速生长的植物。这些黑猩猩的食物有果实、坚果、种子、树叶、蚂蚁、毛虫、蜂蜜和蛋,一旦有机会,它们也食用猴子、丛林野猪以及丛林小鹿的肉。
我们人类是这些现代黑猩猩的后代吗?显然不是,因为并不存在我们可能由之进化的时间。那种想法只是误解而已。事实上,现代人类和现代黑猩猩都是从大约 700 万年前的共同祖先进化而来,那个祖先或许更像黑猩猩而不是人类,因为自那时以后,黑猩猩似乎没有什么变化,而人类以及我们的祖先却发生了巨变。
在 700 万年时间里,自黑猩猩与人类从共同祖先那里分离之后,黑猩猩支系进化出两个物种:普通黑猩猩和倭黑猩猩。人类支系至少出现了 18 种不同物种,其中一些可能一直生活到 5 万年前,尽管只有一个种(智人)存活到了现在(参见图 4.2)。
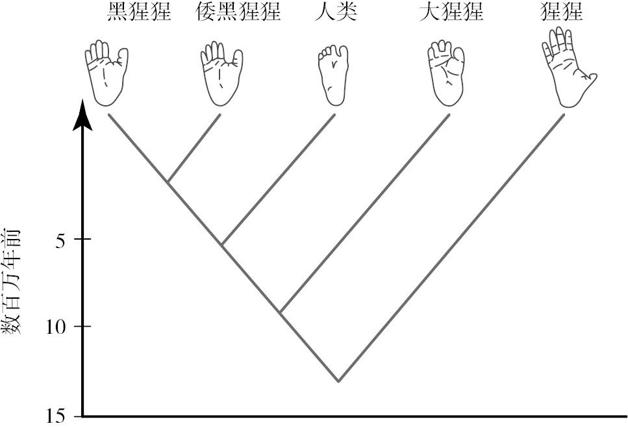
图 4.2 我们的系谱。
你能找出人类与黑猩猩的共同祖先吗?
后面会提到,这两种黑猩猩彼此可能存在很大差异。灵长类动物学家在激烈地争论,到底哪一种更像共同祖先,不过,他们一致认为,与人类相比,这两种动物可能更类似于共同祖先。这个立场的证据在于,人类支系的早期化石很难与现代黑猩猩的化石区分开,而黑猩猩和倭黑猩猩在过去 150 万年变化甚少。一个可能的原因是,这两种黑猩猩生活在雨林,而人类支系则适应了新的环境,即凉爽、干燥气候所创造的开阔的热带稀树大草原。通过研究我们的近亲黑猩猩,我们就能够想象黑猩猩和人类最后的共同祖先是什么模样,只要我们记着他们也发生了某些变化即可。
普通黑猩猩过集群生活,每群 15 到 80 只不等,以具有血缘关系的雄性为中心,每只雄性有众多配偶。雄性确立自己领地的边界,建立边界巡逻兵,并且以武力捍卫自己的领地。雌性通常在青春期之后不久就离开出生的群,加入附近的群;这样就避免了近亲繁殖。人类学家将这种情况称为从夫居(patrilocality),与从母居(matrilocality)相对,后者指雄性移居另一个群。从夫居为雄性提供了血缘优势,而雌性就没有这种亲属支持网络。
雌雄两性都有自己的等级制度,不过,所有雌性都低于雄性。黑猩猩的性关系比较混乱,在配偶和孩子抚养上不存在永久的忠诚,养育幼仔的工作主要由雌性独自承担。多配偶制似乎有利于雌性,因为这样一来,父亲身份就模糊不清了;这正是雌性所想要的,因为一旦雄性黑猩猩知道雄性幼仔不是自己所生,有时会杀死它们。
1960 年之前,人类被认为是唯一使用工具的动物。然而,正是在 1960 年,简·古多尔发现了一个现象:一只黑猩猩把树枝的叶子去掉,然后将它捅进白蚁窝,当树枝被抽出来时,上面沾满了白蚁。此后,研究者观察到,黑猩猩制造和(或)使用了超过半打的工具用于不同目的。每个集群有自己的一套工具,幼年黑猩猩通过观察成年黑猩猩的动作来学习使用这些工具。在整个非洲,黑猩猩集群在使用工具方面各不相同。(现在,我们知道其他一些物种也使用工具,包括一些鸟类和海豚。)
经过 10 年研究,古多尔相信,尽管黑猩猩在行为上非常类似人类,不过,它们大体上更加友好。然而,1971 年,贡贝(Gombe)的研究者观察到了前所未有的行为——对来自相邻集群一只离群雌性的凶残攻击。他们还发现,一位位高权重的雌性与她的女儿(当时没有后代)杀死了自己集群中的许多幼仔。20 世纪 70 年代中期,一群黑猩猩为了拥有自己的领地而从原来的集群中分离出去;此后四年时间,原来的集群对分离群体发动攻击,杀死所有成员(除了三位年轻的雌性),夺回了领地。古多尔只得接受这种结论,即黑猩猩在特定情境中具有强烈的攻击倾向,这些环境包括争夺食物、配偶和领地,或者处于强烈的嫉妒、恐惧和复仇的情绪下。
不过,黑猩猩群体也不断体现出分享、互助、同情以及利他主义。母子和兄弟姐妹之间常常发展出深厚的、一生一世的情谊。兄弟们在成年后成为亲密朋友,在社会冲突中经常同仇敌忾。年长亲属会在年幼黑猩猩丧母后收养它们。非亲属甚至也会收养那些没有年长亲属保护的孤儿。黑猩猩不能游泳;当它们被圈养在四周都是水的动物园时,溺亡事件时有发生。通常情况下,当一只黑猩猩掉进水中,其他的黑猩猩都会冲过去尝试救援。
每一只黑猩猩都可以发出独特的声音,它们拥有丰富的声音、姿势和面部表情,这有利于信息交流。它们的姿势包括伸手讨要食物以及举起双臂要求搭载,9~15 个月大的人类婴儿也经常使用这两个动作。黑猩猩的问候姿势包括拥抱、握手、亲吻和拍打背部。古多尔列举了 34 种不同的发声,它们是咕哝、咆哮、尖叫和叫嚣的不同变化。
黑猩猩的喉——一种由肌肉和软骨组成的结构--位置偏上,有助于防止进食的食道与输送空气的气管交叉。由于这些管道没有交叉,黑猩猩就不会被食物呛到。人类的喉位偏下,这样,我们就获得了一个共振发声的气腔;人类以被呛到的小小风险换来了语言使用的可能。黑猩猩没有共鸣腔和人类那种灵巧的舌头。不过,在人工驯养师的帮助下,黑猩猩能够学会一些基本的符号语言,一些 7 岁大的黑猩猩能够以手势表达 150 个词,沃肖(Washoe,第一只会手语的黑猩猩)能够表达 200~300 个词。黑猩猩可以使用 2~3 个词正确地造句,不过,更长的句子往往有语法错误。(相反,6 岁大孩童掌握的词汇量大约为 1 万个,他们能够说出长句。)在夜间,野生黑猩猩会从树上栖息地不断发出叫声,似乎在确定大家是否安全。即便得到训练,众多黑猩猩也不能同时发声或保持一种节拍。
黑猩猩的另一个种类即倭黑猩猩是在 700 万年前分离出去的,当时,刚果河(1971-1997 年被称为扎伊尔河)阻隔了一群黑猩猩。由于只在群内繁殖,这群黑猩猩发展出了一些明显的特征,这也使得它们变成一个单独的种。今天,它们生活在刚果民主共和国境内,活动范围与英格兰差不多大小。
与黑猩猩较大的胸腔、肩膀和头相比,倭黑猩猩体重比普通黑猩猩稍轻,上身纤细,肩膀较窄,头部较小。倭黑猩猩的腿更长,当站立时,也站得更直;它们更喜欢栖居树上和运动,因为它们生活在浓密而潮湿的沼泽森林地带。潮湿的森林有利于大树生长,这些树木一年四季产出大量果实。由于食物丰富,倭黑猩猩群的数量也更庞大,雌性形成了一种牢固的、安全的纽带关系,尽管它们在成年后会离开最初的集群。一些研究者指出,这也解释了为何倭黑猩猩似乎没有那么刚猛的暴力和攻击性——人们没有观察到强迫性交配、杀婴、边界巡逻队或者对邻居的攻击(参见图 4.3)。

图 4.3 雌性倭黑猩猩(P. panicus)。
据估计,野生倭黑猩猩不足 1 万只
倭黑猩猩有时也会狩猎;雌性通过团结一致实施统治,鼓励集群所有成员之间频繁的、多样化的性活动(异性的和同性的),以此防止大多数攻击行为的爆发。刚果民主共和国爆发内战之后,对野外倭黑猩猩的研究变得困难重重;趁着倭黑猩猩还在野外存在,研究者希望对它们有更多了解。
倭黑猩猩和黑猩猩都以权力和性作为主要关注点。这两个群体对社会地位和其他成员的情感反应表现得很敏感,不断推测谁与谁是同伙。它们的行为在许多方面是人类行为的写照,由此可见,我们的祖先在这些方面肯定与这三个物种都非常相似。
稀树热带草原
在 1500 万年前到 500 万年前,东非的气候发生了剧烈变化。地壳的板块过去和现在继续沿着从莫桑比克到埃塞俄比亚的一条漫长裂缝分离(参见地图 4.1)。非洲东部这块地方最终会与非洲大陆分离,漂浮到印度洋上。同时,这种地壳结构运动导致高地出现,阻断了雨林模式,东非由此拼出不同的栖息地。断裂带两侧的高山和深谷可能隔离了人亚科黑猩猩与它们雨林地区的亲戚。
1000 万年前到 500 万年前,全球气候处于冷却期,650 万年前到 500 万年前,气候尤其严酷。水大量结成冰块,地中海一再干涸。当气候变得更寒冷更干燥时,赤道雨林收缩,外围分裂成许多块林地。历经几百万年的这种气候变化强化了人亚科原人的第一个特征:直立行走。
1994 年,埃塞俄比亚阿法尔三角地带的考古活动取得一项突破性发现:人亚科原人最早种群的骨骼被发现,距今 440 万年。这些不是最后的共同祖先(即露西)的骨骼,不过是迄今为止已知的最接近它的,它们比露西的骨骼早 100 万年,露西的骨骼于 1974 年在离这些骨骼 74 千米以外的地方被发现。这个新发现的种被称为地猿始祖种 (Ardipithecus ramidus),这个名称源自阿法尔语,意思为“地面”和“根”。骨骼被发现的个体被通俗地称作“阿蒂”(Ardi)。
阿蒂是雌性,重 50 公斤,高 122 厘米。她生活在林地,能够直立行走,也能用四肢攀援树枝。其他 36 具骨骼也在附近被发现,雌性和雄性都有较小的犬齿。通过对阿蒂的研究,科学家们断定,最后的共同祖先并不像他们原来所想的那样类似于黑猩猩,直立行走的出现不是由于树木稀缺而是别的原因造成的,小犬齿表明,雄性争夺雌性的冲突减少了,成对结合更常见,这比人们以往所认为的要早。
著名的露西属于南方古猿阿法种,这个种的大多数化石年代介于 350 万年前到 180 万年前。除了一个例外情况,南方古猿化石都来自南部非洲或东部非洲,骨头在那些地方的火山区更容易石化,通过掩埋骨骼的火山灰,也更容易测定它们的年代。不过,史前考古学家认为,南方古猿可能生活在整个非洲地区。它们的特征在于:大脑比黑猩猩的稍大(大约 400~500 立方厘米)、两性异形(雌雄形体大小不一样,雄性比雌性大 50%)以及直立行走。
人类进化为何首先进化出直立行走这个特征呢?当前的解释各种各样。直立行走曾经被认为是为了适应大草原环境,因为那里树木稀少,必须行走才行。然而,1994 年以来,许多早期直立行走的人亚科原人化石在森林地区被发现,这就反驳了早先提出的假说,即直立行走仅仅是为了适应草原环境的需要。当下大量理论认为,黑猩猩发展出直立姿态是为了更有效地从一个林地走到另一个林地,这种姿势使得雄性可以把食物带回给雌性,或者让雄性展示阴茎来吸引配偶。不管这种姿势最初承担的功能是什么,在非洲东部和南部不同居住环境中进化出来的人亚科原人几个种群,因直立行走而获得了繁殖优势。
伴随直立行走而来的,或许是体毛的逐渐消失,尽管没有直接证据证明这一点。毛发减少被认为是在热带大草原保持凉爽的一种适应过程;人亚科原人保留了头顶的毛发,因为头部会受到太阳的直接照射,身体上的长毛逐渐减少,它们还发展出其他纳凉方法,这些都有助于它们在白天采集食物。另一个可能的解释是:雌性会选择毛发较少的雄性,因为它们确信这种雄性身上的寄生虫更少。当然,现代人类也并不是完全没有毛发;我们保留了体毛的痕迹和鸡皮疙瘩。鸡皮疙瘩促使我们毛发竖立,以增加绝缘或作为一种威胁的姿态。
对于早期人亚科原人来说,开阔地带的生活极具挑战性,即便那里还存在少数树木可以用来逃避地面肉食动物的威胁。直立行走是这些早期人亚科原人最早的适应行为,当时还不存在脑容量的快速增长。
灌丛热带大草原
另一段急剧变冷的时期大约开始于 250 万年前,即人属(genus homo)的早期形式出现之际。(这些名称是人类学家起的;Homo 是路易·利基提出来的称呼,他仍然认为制造工具是对人的定义。)这些化石来自脑容量更大、手臂更短、内脏和牙齿更小的生物,这些特征表明,它们吃更多的肉、更少的素食。在这些早期形式中,有鲁道夫人、能人和匠人。这些早期人类——在许多方面类似于类人猿——的头骨与最早的石器工具一起被发现,比如将石块边缘敲掉而形成的简单砍砸器。而那些锐利的石片可能被当作刀片使用。这些石器工具发现于奥杜韦山谷,它们被称为奥杜韦文化(Olduwan);这些工具至少被毫无变化地使用了 100 万年。被称为人(homo)的物种的特征在于:使用工具、离开了树林、脑容量快速发展——在随后 100 万年时间,脑容量从苹果大小扩大到葡萄柚大小。
到 200 万年前,南方古猿和其他各种人属动物以更大的集群生活在开阔地带,同时,它们也食用一些肉(以腐肉为主)。少数人属物种的发声和姿势可能与类人猿的相似,它们使用完整的信息而不是单个词汇影响其他成员的行为。
大约 180 万年前到 170 万年前,一个新物种从多样的人属物种中进化出来,那就是直立人,它们看起来更像人类而不是类人猿。除美洲外,75 具直立人的骨骼已经在世界各地被发现。这个种属到底源自非洲还是亚洲,至今没有定论。一些专家喜欢将早期直立人的骨骼称为匠人。
直立人的骨架几乎与现代人一样高,脑容量是我们的 70% 左右。它们展现出充分的直立行走,双臂较短,不再用来攀援树木。它们的内耳有三个半规管,为跳跃、奔跑和跳舞提供了平衡。与黑猩猩相比,它们(不管雄性还是雌性)的骨盆变得更窄、更扁平,这为充分站立和奔跑奠定了基础,也缩短了盆骨内产道的直径。
对前面所说的两类黑猩猩而言,生育很轻松,雌性能够独自完成这项工作。但对直立人来说,产道变小了,幼儿的头却增大了,这使得生育变得更困难。这样,幼儿就得更早出生(事实上没有发育成熟),即在头部增长得太大之前就出来。一个现代婴儿的大脑在出生时只是成年人的 23%,黑猩猩的是 45%。直立人的盆骨表明,雌性无法生育出大脑为成人大脑 45% 的幼儿;因此,出现了出生后大脑快速发展的模式。
更多无助的新生儿需要更长期和更精细的照料;雌性需要雄性的帮助,以便喂养幼仔以及保护母亲和幼儿免遭其他雌性和肉食动物的伤害。人亚科原人以某种方式改变了黑猩猩那种非长久的雌雄结合模式,在儿女的父母之间发展出了一种至少暂时性的配偶关系。为了在照看孩子上获得保护和帮助,或许雌性和雄性都放弃了性自由,以便增加它们后代的存活机会。不管怎样,直立人发展出了持久的合作和互助模式。
成对结合的间接证据在于,这一时期雄性骨骼比雌性的只大 25%,而南方古猿的这一数据是 50%。两性异形的缩减表明雄性争夺雌性的竞争变得更少。(现代男性体型平均只比女性大 15%~20%。)
除了成对结合之外,直立人可能也学会了使用火,它们用火烹调和保存食物——或许包含了高比例的肉以及一些块茎或根。熟食减少了撕咬和咀嚼的时间,肠道也可以缩短,这很可能是大脑扩大的前提条件,理查德·兰厄姆(Richard Wrangham)在他的著作《生火:烹饪如何造就人类》(2009)中就是这么认为的。
火的使用是人亚科原人迈出的最重要步伐之一;它也是把我们与其他动物区分开的一个特征,并且为随后的各种发展奠定了基础。围绕火展开的社会活动领域可能促进了语言的发展,也推动了更高层次的工具制造技术,即阿舍利文化(the Acheulian),其典型工具是大型的、刃缘规整的双刃手斧,这种工具至少持续使用了 100 万年(一直用到 25 万年前)。火的使用增加了能量在人亚科原人社群中的流动。
直立人是第一个走出非洲的人亚科种群,这也使得它与众不同,尽管最近的研究指出,做到这一点的,很可能是更早的亲属——匠人。直立人的一些集群至少在 180 万年前来到欧洲边缘高加索地区的格鲁吉亚。在冰河期环境下,它们在那里进化为尼安德特人,即智人的近亲,一直生活到大约 3 万年前到 2 万年前。存在多少交流呢?没有人知道答案。主流观点认为,直立人说一种原始语言,使用简单名词和动词,这种语言在语速、词汇和复杂性方面逐渐得到增强。
在直立人生活时期,另一个可能出现的变化是肤色变深。黑猩猩的黑色毛发掩盖着浅色皮肤。当早期人亚科原人失去它们的毛发时,苍白的皮肤致使它们很容易遭到紫外线辐射的伤害,导致皮肤癌和一种叫叶酸的重要营养物质被摧毁,从而降低生育能力。遗传学家认为,到 120 万年前,有利于深色皮肤的基因突变可能席卷整个非洲。在非洲之外地区,早期人类的皮肤颜色再次变浅,以便帮助它们合成足够多的维生素 D,而深色皮肤一定程度上会妨碍这种物质的合成。在当前世界每一个民族中,女性皮肤比男性浅 3%~4%,没有人知道为何如此。这种现象的原因到底是性别选择,还是出于对维生素 D 的更多需要,抑或其他因素呢?
最近发现了直立人的两条支脉。其中之一是鲁道夫人的骨骼,于 2003 年在爪哇东部一个海岛发现。所发现的 9 具不完整骨骼属于人亚科一个种群,身高 1 米多一点,通常被称为“霍比特人”。它们或许是直立人,或许不是。2010 年,在俄国、蒙古、中国和哈萨克斯坦交界处阿尔泰山地区的丹尼索瓦洞(Denisova Cave)发现了一些手指和脚趾骨骼。这些丹尼索瓦人距今 41000 年,与尼安德特人和现代人不同。
我们对人亚科原人的进化做了简要考察,描述了黑猩猩和人类共同祖先的各种特征如何演化为现代人的特征。在大约 700 万年时间里,脊骨变得更直、骨盆更窄、脑容量更大、手臂更短、成对结合、交流和合作增强、火得到利用、手斧得以铸造——所有这些都发生在气候急剧变化的环境中。
直立人的后代散布到整个欧亚大陆,并且以几种不同的方式演化。在亚洲,它们变化很少,还是同一种群;在欧洲,它们适应了冰期环境,转变为尼安德特人;在非洲,它们进化为现代人(智人)。尼安德特人是否是不同于智人的单独种群,它们在何种程度上是智人的祖先,这些问题一直备受争议。当尼安德特人生活在欧洲和中东时,智人正在非洲进化,因此,接下来我们会考察智人。